Le Tissu Conjonctif (cours N°1, 2008)Le Tissu Conjonctif
A) Généralités : Le tissu conjonctif non spécialisé (nommé tissu conjonctif) est présent dans la totalité des organes, avec toutefois des variations qualitatives et quantitatives majeures dun organe à lautre. Le cerveau par exemple est très pauvre en tissu conjonctif alors que le muscle est riche en TC. Le terme de tissu conjonctif désigne les tissus conjonctifs communs, non spécialisés. Il ninclut pas les tissus conjonctifs spécialisés cest-à-dire : le tissu adipeux, le tissu osseux et le tissu cartilagineux. On le verra plus loin, il existe néamoins différentes catégories de tissu conjonctif commune, non spécialisé. La matrice extra-cellulaire du TC est caractérisée par la présence de molécules fibreuses qui sont synthétisées par les cellules résidentes du tissu conjonctif : les fibroblastes. Selon sa localisation et son sous-type, le tissu conjonctif assure 3 fonctions principales : i) une fonction de soutien et de cohésion, liée à sa richesse en fibres ;
ii) une fonction nutritive et déchange, liée à sa richesse en matrice extracellulaire et en vaisseaux ;
iii) une fonction de défense, liée à la présence de cellules immune dites de passage.
Le terme de tissu conjonctif désigne les tissus conjonctifs communs, non spécialisés. Il ninclut pas le tissu adipeux, le tissu osseux et le tissu cartilagineux.
B) la matrice extra-cellulaire du tissu conjonctif
1) Notions générales sur les matrices extra-cellulaires : Tous les tissus sont formés par lassociation de cellules et dun milieu extra-cellulaire nommé matrice extra-cellulaire (MEC). Selon le tissu considéré, la MEC présente dimportantes variations quantitatives et qualitatives. Le tissu le plus riche en MEC est le tissu conjonctif et celui le plus pauvre en MEC est le tissu épithélial (La MEC y est en effet limitée à la matrice des espaces para-cellulaires et à la lame basale). Les MEC sont constituées principalement de 3 éléments : i) des fibres formées de lassemblage de protéines fibreuses dites de structure. Ces fibres ne sont présentes que dans la MEC des tissus conjonctifs et dans la MEC formant les lames basales. Elles sont visibles en microscopie électronique.
ii) de leau
iii) des molécules solubles tels que des sels minéraux, des polypeptides des sucres et des macromolécules protéiques. Leau et les molécules solubles forment la substance fondamentale. Celle-ci est dite optiquement vide i.e non visible en microscopie électronique.
2) Les molécules de la matrice : Les molécules participant à la MEC du tissu conjonctif peuvent être observées, pour partie ou en totalité, dans tous les types de matrice. Elles se divisent en 3 catégories : les polysaccharides, les protéines dadhérence et les protéines de structure. Seules les protéines de structure ont la capacité de former des fibres, les autres molécules (polysaccharides et molécules dadhérence appartiennent à la substance fondamentale.
a) polysaccharides ils sont répartis en deux familles.
* Les glycosaminoglycanes sont soit sulfatés (pour mémoire : chondroïtine-sulfate, le dermatane-sulfate, l'héparane-sulfate, le kératane-sulfate) soit non sulfatés (l'acide hyaluronique). Lacide hyaluronique présente la caractéristique de pouvoir lier de nombreuses protéines de la MEC du tissu conjonctif (pour mémoire : protéine fibreuses telle que le collagène, ou protéines d'adhérence telles que la fibronectine et la laminine). Lacide hyaluronique est également un ligand pour des récepteurs membranaires qui, dans le tissu conjonctif, sont exprimés par les fibroblastes et/ou par les cellules immunes de passage. Il sagit en particulier de la molécule CD44 et des molécules de la famille Toll-like Receptor (TLR). On considère que linteraction entre acide hyaluronique et ses récepteurs membranaires joue un rôle essentiel dans les processus dinflammation et de réparation tissulaire, au sein du tissu conjonctif (Rappel : dans les tissus épithéliaux, l'acide hyaluronique présent dans la lame basale interagit avec la molécule CD44 exprimée par les cellules épithéliales).
* Les protéoglycanes sont formés par un noyau protéique sur lequel se lient des glycosaminoglycanes sulfatés. Ces protéoglycanes (pour mémoire : syndecan, perlecan) forment des aggrégats de très grande taille dont le squelette est formé de molécules d'acide hyaluronique et qui ont la capacité de fixer certaines cytokines ou facteurs de croissance et donc de moduler leur biodisponibilité (pour mémoire: les aggrégats de protéoglycanes s'articulent autour de molécules d'acide hyaluronique).
 b) les protéines d'adhérence : comme les polysaccharides, ce sont des molécules solubles entrant dans la composition de la substance fondamentale.
* la fibronectine :
* la laminine : il s'agit d'une protéine d'adhérence présente dans les tissus conjonctif bien quessentiellement localisée au niveau des lames basales
mais qui est prédominante au niveau des lames basales et non du TC. Elle lie certains polysaccharides tel que l'acide hyaluronique ainsi que les récepteurs membranaires de la famille des intégrines.
c) les protéines de structure :
* le collagène : cest la plus abondante des protéines de lorganisme et on considère quelle constitue un quart du poids sec des mammifères. On reconnaît actuellement plus de 20 types de molécules de collagène. Le collagène I est le plus abondamment distribué dans le tissu conjonctif. Les molécules de collagène I forment des microfibrilles visibles en microscopie électronique et présentant une striation transversale due à l'alternance de bandes sombres et claires.
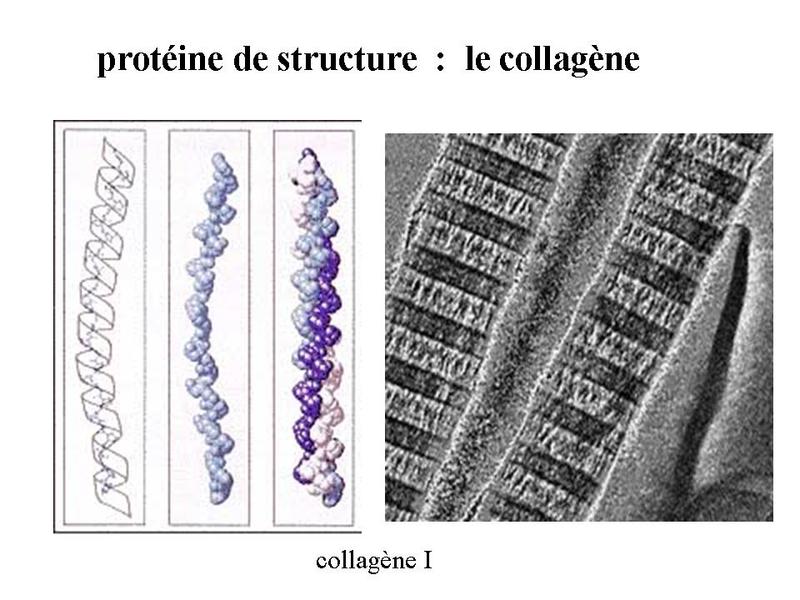 Ces microfibrilles se groupent pour former des fibres qui s'assemblent en faisceaux diversement orientés dans lespace . Cette organisation confère des propriétés de soutien mécanique au tissu conjonctif. Ces fibres et faisceaux sont visibles en MO après coloration au jaune safran ou au trichrome de Masson.
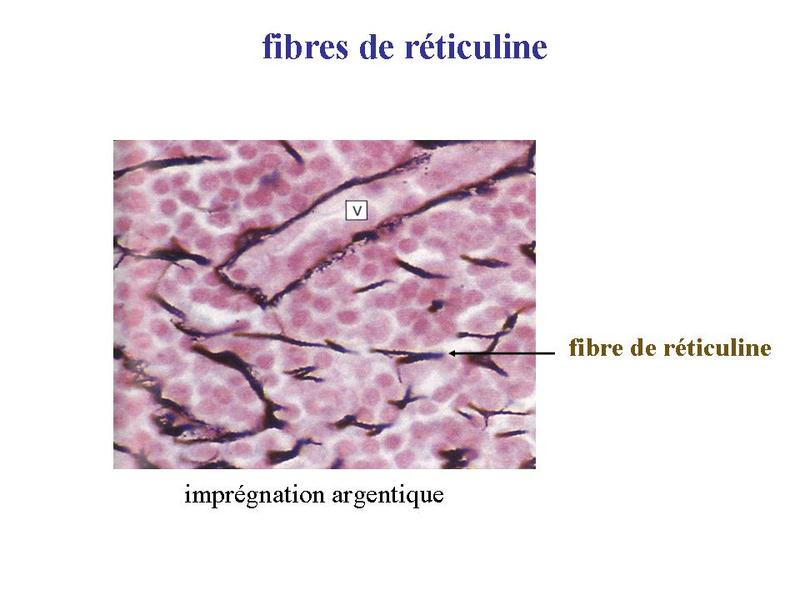
 Une catégorie de tissu conjonctif, le tissu conjonctif réticulaire, contient majoritairement du collagène de type III et non de type I ; Le collagène type III forme des fibres dites de réticuline qui ne sont pas visualisables par le trichrome de Masson ou le jaune safran.
 Par ailleurs, il faut retenir que le collagène de type II est caractéristique du tissu cartilagineux et que le collagène de type IV est essentiellement présent au niveau des membranes basales.
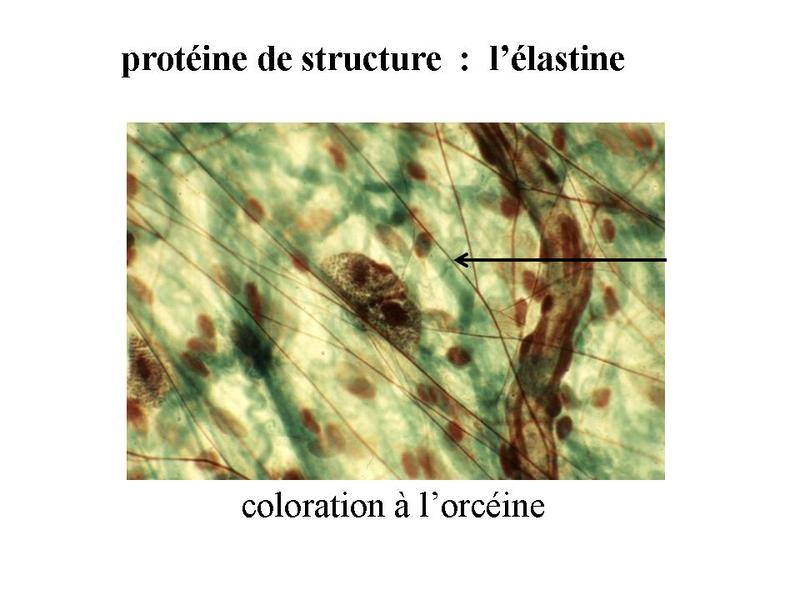
* lélastine : cest la molécule principale des fibres élastiques qui sont présentes dans les tissus conjonctifs élastiques. En MO, les fibres élastiques sont visibles par la coloration à lorcéine sous forme d'un réseau de fines fibres allongées et anastomosées.
 C) Cellules du tissu conjonctif :
1) fibroblastes : le fibroblaste est le seul type cellulaire présent dans tous les tissus conjonctifs (TC communs, non spécialisés). Il s'agit d'une cellule fusiforme, porteuse de prolongements plus ou moins ramifiés. En microscopie électronique, le cytoplasme des fibroblastes est riche en organites impliqués dans la synthèse protéique : il s'agit d'une cllule sécrétrice de protéines. Les fibroblastes synthétisent la matrice du tissu conjonctif et sont le plus souvent entourés des protéines fibreuses dont ils sécrètent les molécules précurseurs. Cest le cas notamment du pro-collagène et de la pro-élastine qui sont les formes non fibrillaires du collagène et de lélastine.
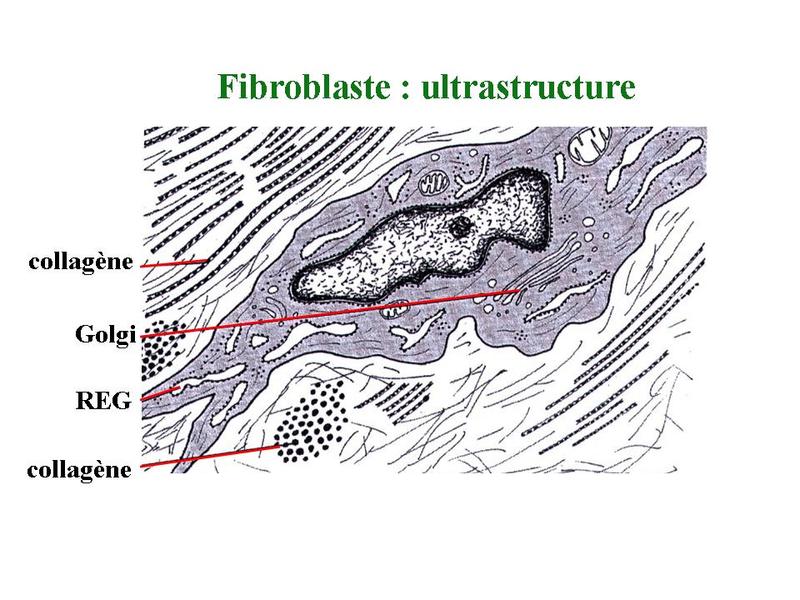 Le terme de fibrocyte désigne des fibroblastes en fin de vie qui ne sont plus capables de division cellulaire et synthétisent peu de matrice. Les fibroblastes dérivent de cellules souches mésenchymateuses. Ces cellules sont à lorigine de tous les types cellulaires spécifiques des tissu conjonctifs spécialisés ou non spécialisés : les fibroblastes (qui se différencient en fibrocytes), les adipoblastes (qui donnent les adipocytes), les chondroblastes (qui génèrent chondrocytes) et les ostéoblastes (a lorigine des ostéocytes). Chez l'adulte, des travaux récents montrent que les cellules souches mésenchymateuses i) persistent dans les tissus conjonctifs non spécialisés (dans le tissu conjonctif associé à la moelle osseuse par exemple) ii) présentent un potentiel de différentiation plus large quinitialement observés (potentiel de différentiation neuronale par exemple). Par ailleurs, certains auteurs ont récemment mis en évidence l'existence de précurseurs sanguins des fibroblastes, précurseurs qui seraient recrutés sur les sites inflammatoires et participeraient à la reconstruction tissulaire ou à la formation de lésions cicatricielles fibrotiques (fibros cicatricielle).
La synthèse protéique des fibroblastes ne se limite pas à la sécrétion de molécules de la matrice. Outre les molécules de la MEC. Les fibroblastes synthétisent des cytokines et notamment de linterféron-beta qui joue un rôle important dans les défenses anti-virales. Ils synthétisent également des cytokines pro-inflammatoires et des chimiokines qui participent au micro-environnement des tumeurs. Le rôle permissif de ce micro-environnement inflammatoire est fortement suspecté (facilitation de linvasion et de la dissémination métastatique).
2) adipocytes : ce sont des cellules spécialisées dans la mise en réserve des lipides. Les adipocytes sont regroupées en amas au sein de la plupart des tissus conjonctifs non spécialisés. Le terme de tissu adipeux désigne un tissu conjonctif spécifique au sein duquel les adipocytes sont très largement majoritaires. Les adipocytes sont de morphologie sphérique ou polyédrique avec un cytoplasme essentiellement occupé par une vacuole lipidique et un petit noyau refoulé contre la membrane plasmique. Les adipocytes ne sont pas de simples cellules de stockage lipidique.
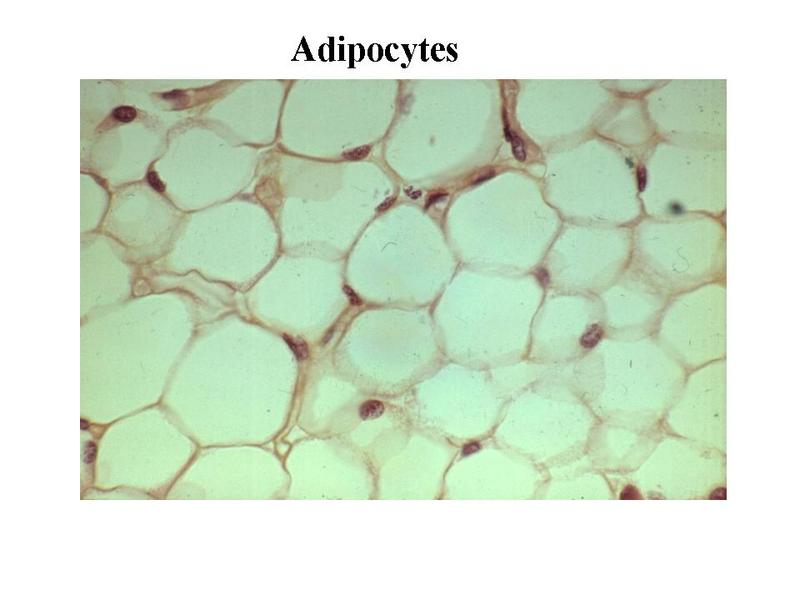 Les adipocytes synthétisent un grand nombre de molécules contrôlant le métabolisme lipidique, la prise alimentaire et la réponse immune. Certains auteurs parlent dadipokines pour désigner ces molécules. Ainsi, la leptine, synthétisée par les adipocytes, est relarguée dans le sang et agit au niveau de récepteurs hypothalamiques pour réguler la sensation de satiété. De la même façon, les adipocytes sont une importante source de TNF-alpha (tumor necrosis factor-alpha), cytokine pro-inflammatoire impliquée dans lamaigrissement prononcé observé au cours des stades terminaux du cancer. Laugmentation massive du volume du tissu adipeux est une caractéristique de lobésité sévère (encore appelée obésité morbide). On a longtemps considéré que les adipocytes pouvaient être soumis à des variations de volume mais non de nombre. En fait, au cours des obésités sévères (obésités dites « morbides »), il existe une augmentation du nombre dadipocyte qui serait lié à une différenciation adipocytaire de cellules souches mésenchymateuses résidant dans le tissu adipeux. Par ailleurs, lobésité sévère saccompagne dune infiltration du tissu adipeux par des macrophages et par une altération du statut immunitaire des patients.
3) cellules immunes : Les tissus conjonctifs dits lâches, c'est-à-dire riches en cellules et pauvres en fibres (Cf infra), sont dimportants lieux de transit pour les cellules immunes sanguines. Selon la localisation du tissu conjonctif lâche, on observe des variations qualitatives et quantitatives de ces populations cellulaires immunes. Ces variations sont corrélées à des statuts immunitaires différents. Par exemple, le chorion de la muqueuse intestinale (c'est-à-dire le tissu conjonctif sous-jacent à lépithélium intestinal) contient de nombreux lymphocytes T et des plasmocytes sécrétant des Immunoglobulines de type A. Par comparaison, le derme (le tissu conjonctif sous-jacent à lépiderme) contient des cellules dendritiques (les cellules dendritiques du derme sont à distinguer des cellules de Langerhans de lépiderme), des lymphocytes T mais pas de plasmocytes .
a) les macrophages : ils dérivent de monocytes sanguins ayant pénétrés dans les tissus conjonctifs lâches. Ils présentent 4 caractéristiques morphologiques principales :
i) il sagit de cellules rondes ou ovalaires de très grande taille (diamètre de 50 micron ou plus alors que les monocytes ont un diamètre inférieur à 20 microns)
ii) leur noyau est excentré et présente une morphologie réniforme ou encochée
iii) lappareil vacuolaire y est très développé et comprend des vésicules de pinocytose, des lysosomes primaires, des phagosomes et des phagolysosomes
iv) on observe des expansions de la membrane plasmique formant des pseudopodes. Ces pseudopodes sont nécessaires à la mobilité des macrophages.
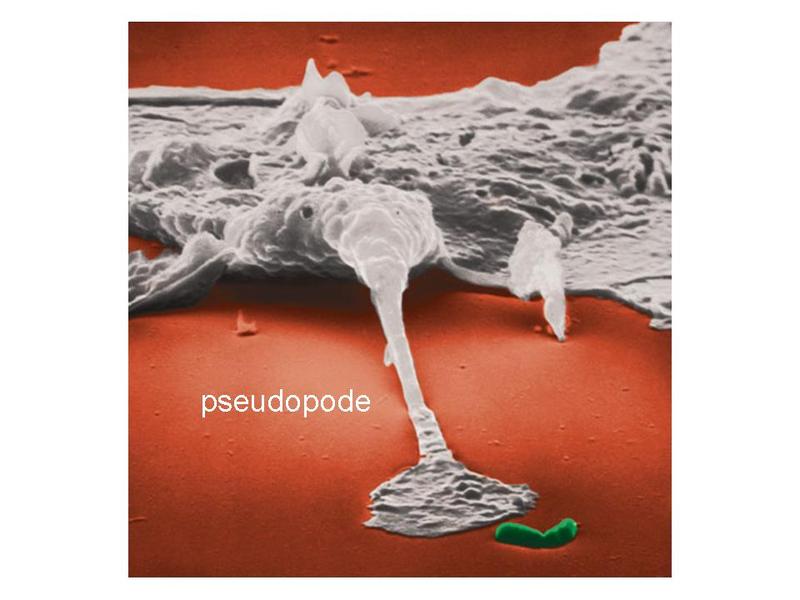  Les macrophages exercent principalement 3 fonctions. La première est la phagocytose. Elle sexerce vis-à-vis de micro-organismes, de débris cellulaires, de cellules en apoptose et dantigènes associés à des anticorps (complexes antigène/anticorps). Cette activité met en jeu un ensemble de récepteurs membranaires capables de reconnaître spécifiquement les cibles à phagocyter. Par ailleurs, selon le recepteur membranaire impliqué, la fonction de phagocytose est couplée à une activation cellulaire dont les caractéristiques sont spécifiques du récepteur. La deuxième fonction des macrophages est la présentation dantigènes aux lymphocytes T. Elle nécessite une phase dite dapprêtement de lantigène. Il s agit dune digestion sélective de lantigène conduisant à la formation de multiples peptides qui seront présentés à la surface membranaire en association avec les molécules dhistocompatibilité. Cette phase dapprêtement est accompagnée dune phase de migration des macrophages depuis le TC jusquaux ganglions lymphatiques ou seffectue linteraction avec le lymphocyte T. La troisième fonction est la synthèse de cytokines pro-inflammatoires permettant lamplification de la réponse immune. Cest le cas notamment de lIL-1 (interleukine-1), de l'IL-6 (interleukine-6) et du TNF-alpha. Il faut noter toutefois que certains signaux dactivation des macrophages conduisent au contraire à la synthèse de cytokines dites anti-inflammatoires (pour mémoire : linterleukine 10). b) les plasmocytes : les plasmocytes correspondent au stade ultime de différenciation des lymphocytes B et sont les seules cellules du système immunitaire capables de sécréter des immunoglobulines. Il sagit de cellules ovoïdes dont le noyau excentré présente une chromatine dite « en rayons de roue ». Les plasmocytes possèdent un volumineux appareil de Golgi et un réticulum endoplasmique granuleux abondant.
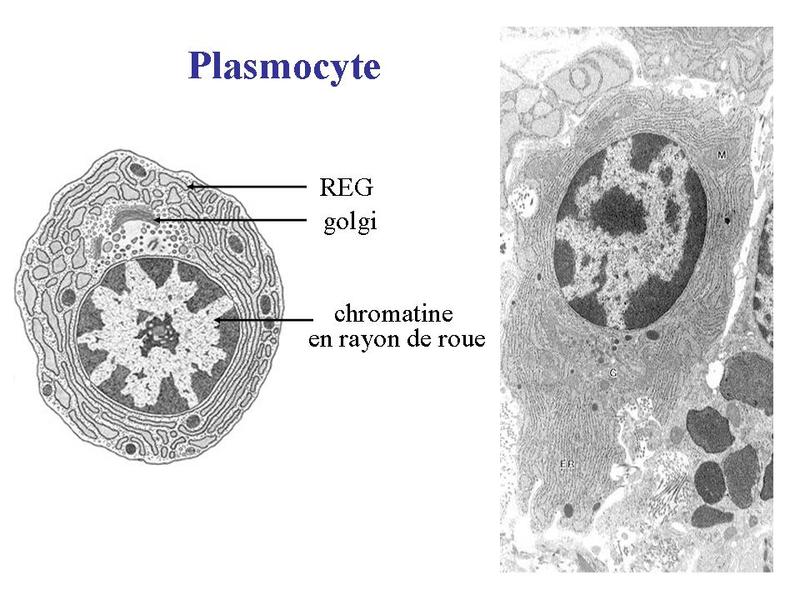 Les plasmocytes sont des cellules non circulantes (cest-à-dire non présentes dans le sang), présentes exclusivement au niveau des tissu conjonctifs lâches, des organes lymphoïdes secondaires (ganglions, rate) et de la moelle osseuse (organe lymphoïde primaire). c) les mastocytes : ce sont des cellules arrondies, à noyau central, et caractérisées par la présence dabondants grains de sécrétions intracytoplasmiques. Ces grains (encore nommés granulations) contiennent essentiellement de lhistamine qui est une molécule fortement vaso-active.
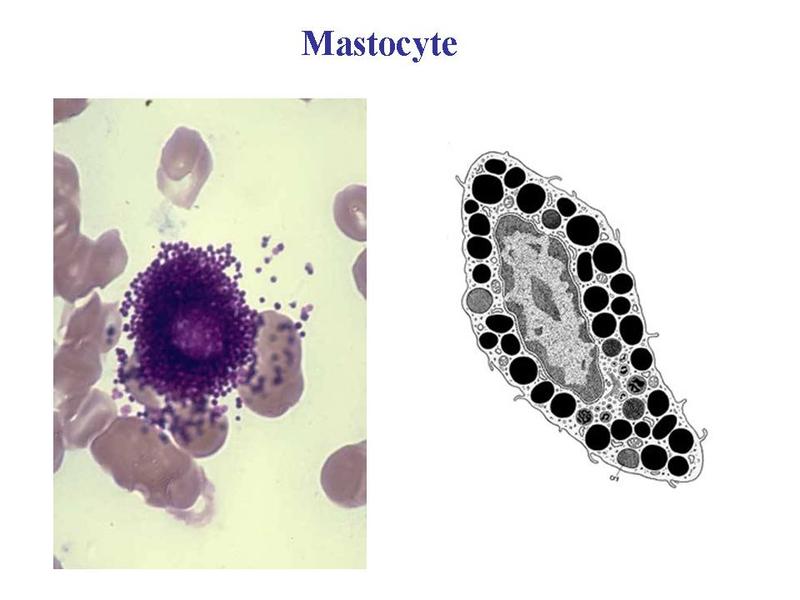 La dégranulation des mastocytes cest-à-dire la libération du contenu des granulations, par exocytose, est induite principalement par la fixation dimmunoglobulines de type E (Ig E) à des récepteurs membranaires spécifiques (recepteuirs aux IgE). Elle intervient lors de phénomènes allergiques aigus en induisant une vasodilation locale avec infiltration massive de cellules immunes sanguines au sein du tissu conjonctif. 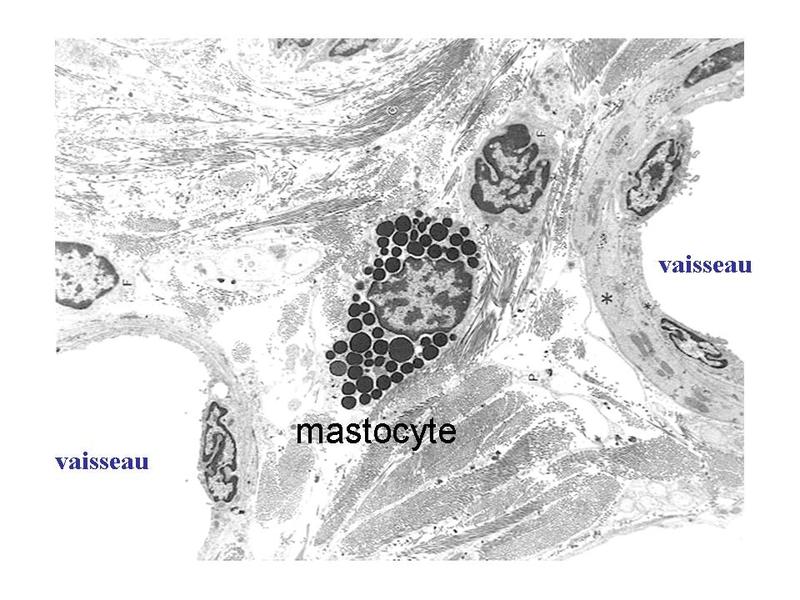 Les mastocytes sont particulièrement abondants dans le tissu conjonctif de la peau (derme, hypoderme), des bronches et des voies digestives. Cette localisation préférentielle explique leur rôle dans la physiopathologie de maladies allergiques cutanées (eczema, urticaire), pulmonaires (asthme) ou digestives (allergies alimentaires). Dans ces pathologies, les traitements antihistaminiques sont largement utilisés. D) Classification
Il existe 4 catégories de TC non spécialisé : le TC lâche, le tissu conjonctif réticulaire, le TC dense et le TC élastique.La classification des tissus conjonctifs non spécialisés
est basée sur 2 critères : i) la richesse relative en fibres par rapport à la richesse en cellules et en substance fondamentale ; ii) la nature des fibres.
1) le tissu conjonctif lâche :
a) généralités : Cest le tissu conjonctif le plus répandu dans lorganisme.
Il est relativement pauvre en fibres comparés aux tissus conjonctifs denses. En revanche il est riche en cellules et/ou en substance fondamentale. Par ailleurs il est richement vascularisé.
Il exerce principalement des fonctions de défense immune, de soutien nutritif et métabolique et de soutien mécanique. On retient 4 principales catégories de TC lâche :
i) les TC sous-jacents aux épithéliums de revêtement i.e ; le derme et lhypoderme au niveau de la peau et le chorion au niveau des muqueuses ;
ii) le TC de la paroi des veines et des artères de faible calibre;
iii) le tissu conjonctif associé aux nerfs périphériques et aux muscles ;
iv) le TC du parenchyme des organes pleins et qui est nommé stroma conjonctif.
Tous les organes pleins comme le rein le foie ou le pancréas contiennent un abondant tissu conjonctif qui rempli les interstices entre cellules et assure la cohésion de lorgane. On peut dire que le parenchyme des organes pleins est constitué de cellules spécialisées et dun stroma conjonctif. Une exception à cette règle : le parenchyme du système nerveux central. A retenir toutefois quon trouve du TC dans le système nerveux central, au niveau de la paroi des vaisseaux sanguins non capillaires. |
Site créé avec ViaBloga
-- Modèle rdc par Jon Roobottom,
Paul Lloyd et
Stéphane Gigandet --
![]()