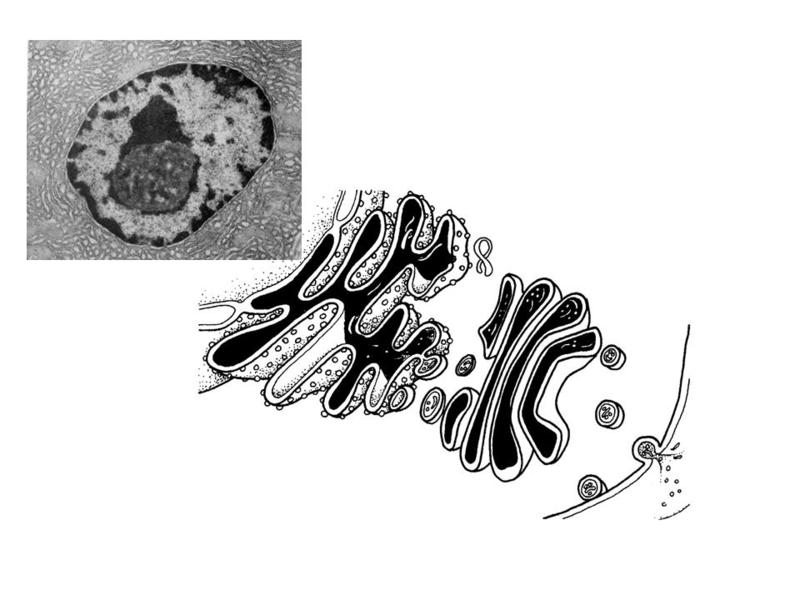
Le Tissu Epithélial (cours N°3)IV) EPITHÉLIUMS GLANDULAIRES A) Cellules sécrétrices : Les cellules de lorganisme ont pratiquement toutes une activité sécrétrice caractérisée par la synthèse puis la libération dune molécule biologiquement active. Toutefois, il existe des cellules spécialisées dans la sécrétion. Ces cellules présentent une morphologie et une cytophysiologie qui dépendent principalement du type de molécule sécrétée. Il existe essentiellement 3 grandes catégories de molécules sécrétées : les protéines, les hormones et les neuromédiateurs 1) les protéines : a) généralités : dans les cellules à faible activité sécrétrice, la synthèse et la libération de protéines seffectuent sur un mode continu et non régulé (la sécrétion est donc constante et maintenue à une niveau relativement faible). Dans les cellules à forte activité sécrétrice, la sécrétion est quantitativement variable et régulée par des signaux extrinsèques. Au plan morphologique, les cellules présentant une forte activité de sécrétion protéique sont caractérisées par : un nucléole volumineux, un REG et un appareil de Golgi très développé, la présence de vésicules (ou grains) de sécrétion. La libération du contenu des vésicules seffectue par exocytose (fusion entre la membrane vésiculaire et la membrane plasmique).
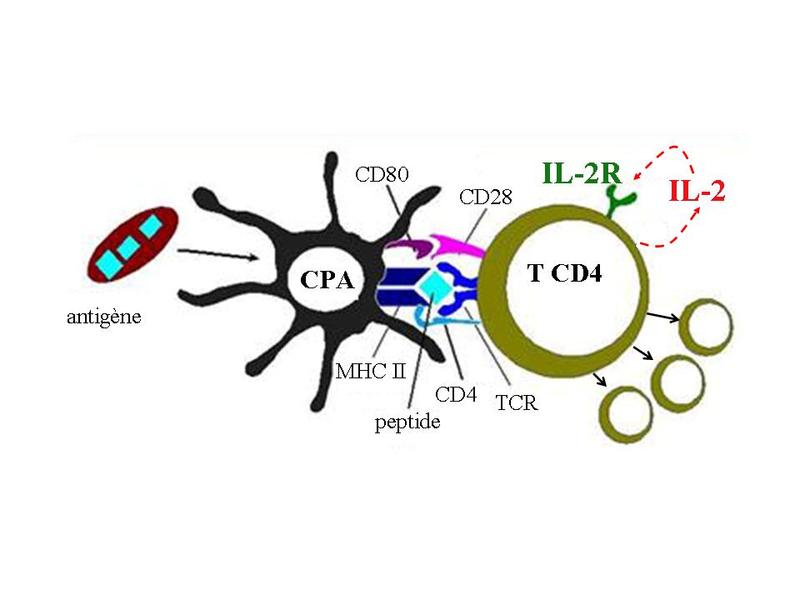
Dans la catégorie des protéines sécrétées on trouve notamment lensemble des molécules de la matrice extracellulaire, les facteurs de croissance, les protéines de sécrétions muqueuses, les enzymes, les cytokines, les immunoglobulines etc.. Nous détaillerons rapidement lune de ces familles de molécules : les cytokines. b) les cytokines : Le terme de cytokine désigne principalement les protéines sécrétées présentant une activité immunologique. Les premières cytokines identifiées ont été les interleukines ( assurant la communication « entre leukocytes » cest-à-dire les globules blancs) et notamment linterleukine-1 qui est synthétisée par les monocytes/macrophages et linterleukine-2 qui est synthétisée par les lymphocytes T. Les cytokines ne sont pas nécessairement synthétisées par des cellules immunes et leurs récepteurs membranaires nont pas une expression restreinte aux cellules immunes. Se rapproche des cytokines, une famille de molécules très actives au sein du système immunitaire : les chimiokines. Il sagit de molécules capables dinduire le déplacement de cellules à travers les tissus. Elles agissent par fixation sur des récepteurs membranaires spécifiques qui sont des récepteurs à 7 domaines trans-membranaires couplés à une protéine G. On dit que la cellule se déplace contre un gradient de chimiokine.
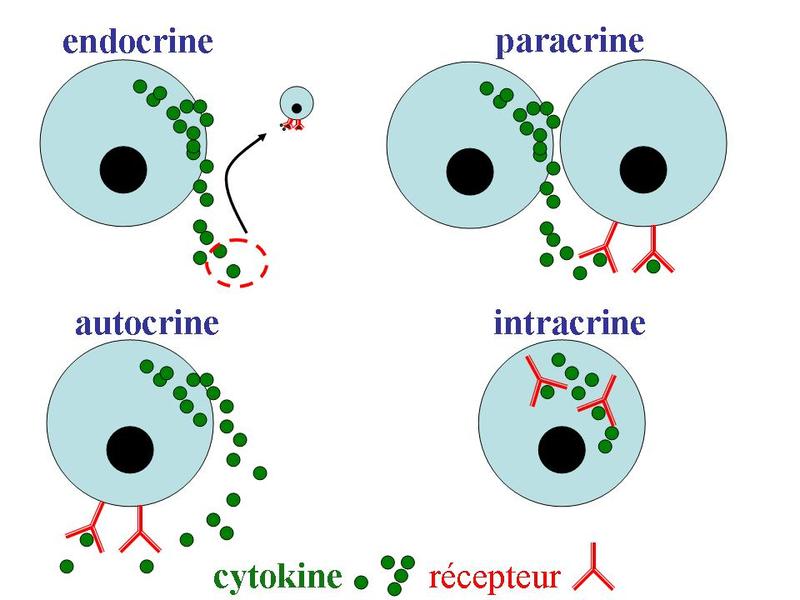
Les cytokines présentent 4 grandes modalités daction selon la localisation de leur cible cellulaire : - on parle de modalité endocrine lorsque la cytokine est sécrétée dans le sang et agit sur un site daction localisé à distance du site de sécrétion : cest le cas de linterleukine-1 qui est synthétisée massivement par les cellules immunes en situation dinfection et qui est responsable, par voie endocrine, de perturbations du fonctionnement cérébral (perte dappétit, somnolence, fièvre). - on parle de modalité paracrine lorsque la cellule cible est adjacente à la cellule sécrétrice. Par exemple, linterféron-gamma synthétisé par les lymphocytes T CD4 activés induit lactivation des macrophages adjacents à la cellule sécrétrice. - on parle de modalité autocrine lorsque la cellules cible est la cellule sécrétrice de cytokine. Par exemple, les lymphocyte T CD4 activés synthétisent de linterleukine-2 qui induit leur prolifération.
- on parle de modalité intracrine, lorsque la cytokine est sécrétée à lintérieur même du cytoplasme de la cellule. Cest le cas de linterleukine-1 qui peut agir sur le mode intracrine en modulant lactivité transcriptionnelle de la cellule sécrétrice. Ce mode daction est très rare.
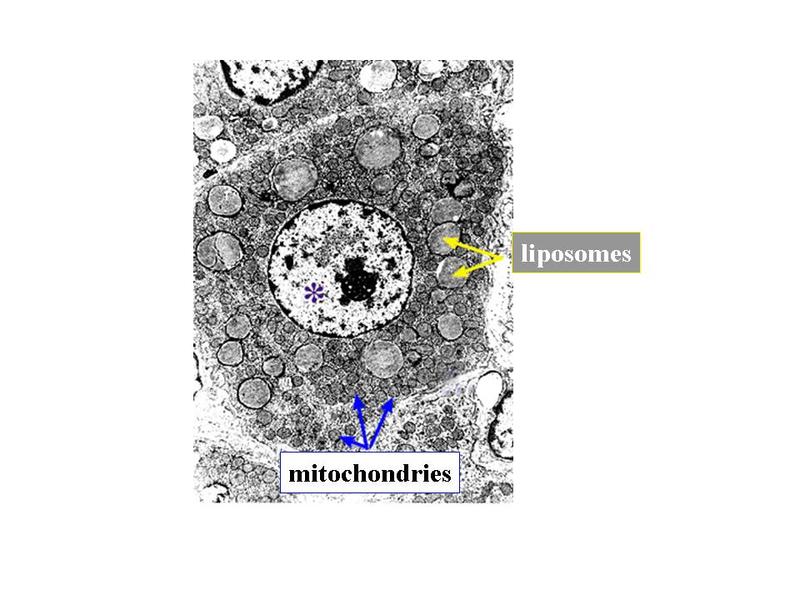
2) les hormones : Les cellules sécrétrices dhormones (au sens strict du terme) appartiennent au système endocrinien et sont nommées cellules endocrines. Un exemple est donné par les cellules sécrétrices dhormones stéroïdes telles que les minéralocorticoïdes, les glucocorticoïdes, les androgènes et les oestrogènes. Ces cellules qui sont localisées dans la glande surrénale et dans les gonades présentent des caractéristiques morphologiques communes : i) la richesse en REL, ii) labondance de mitochondries avec la particularité que ces mitochondries possèdent des crêtes tubulaires et non lamellaires, iii)) la présence de liposomes cest-à-dire de vacuole contenant un matériel lipidique.
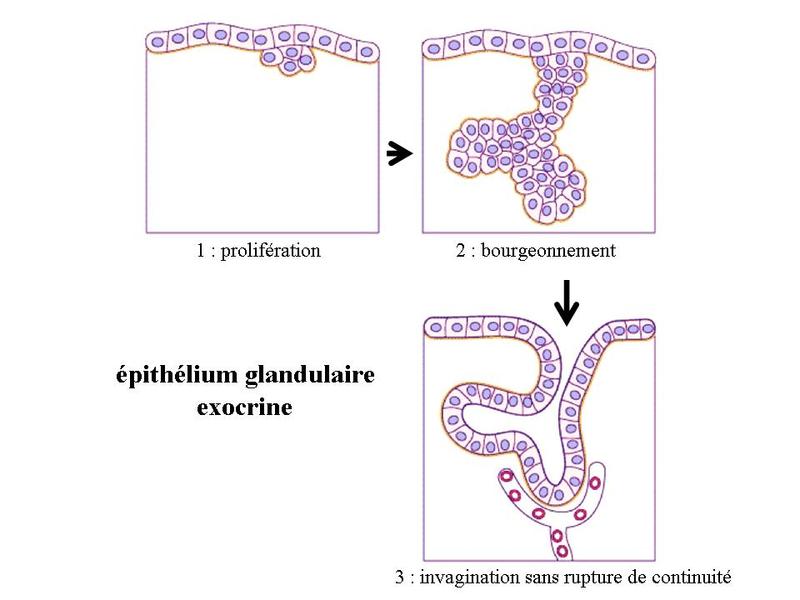
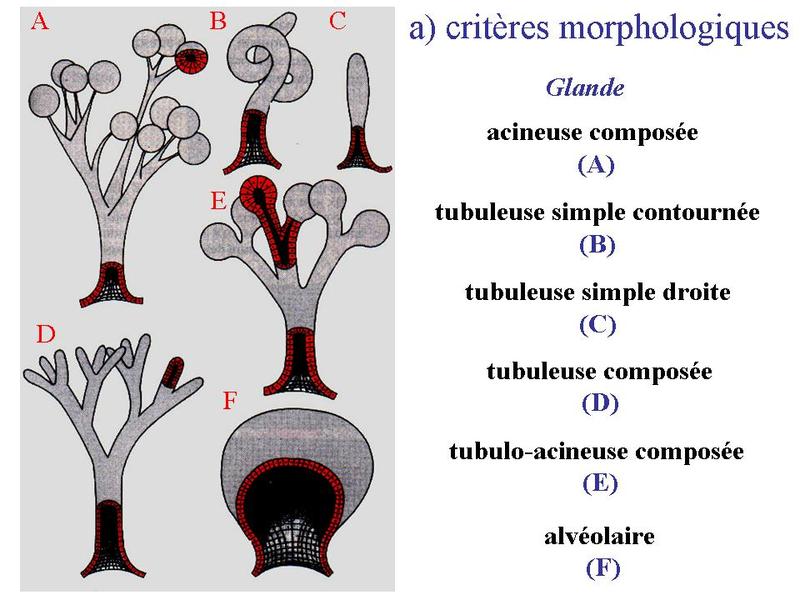
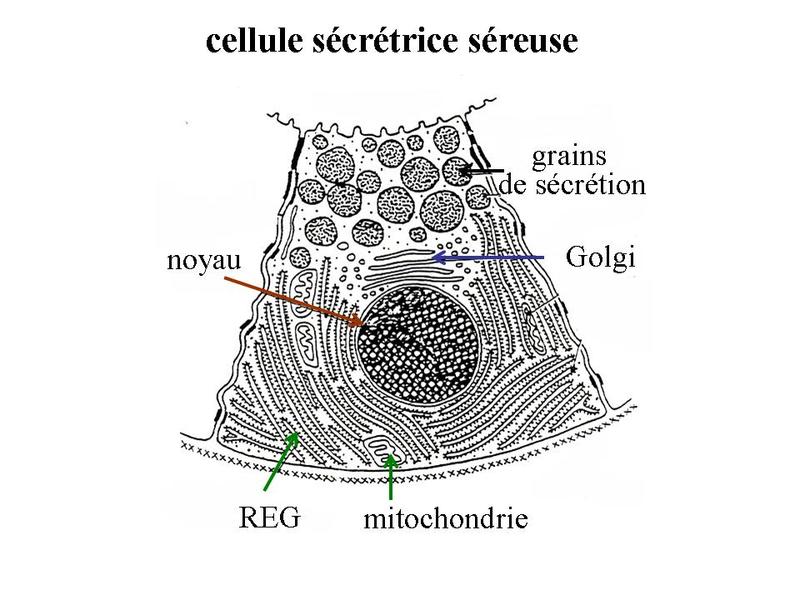
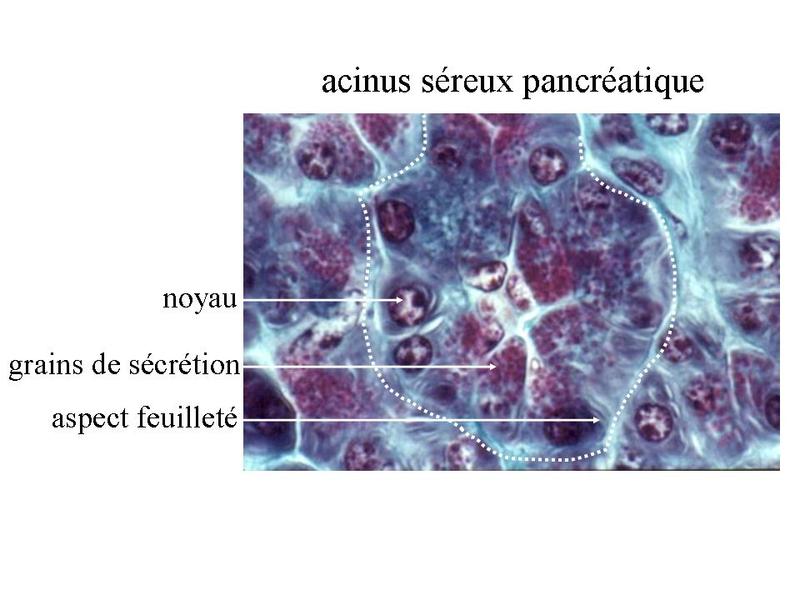
Ces particularités ultrastructurales sont à mettre en relation avec le mode de synthèse des hormones stéroïdes. Cette synthèse seffectue à partir de cholesterol apporté par lalimentation ou synthétisé par la cellule et stocké au sein des liposomes. Le cholestérol est ensuite métabolisé en hormones stéroïdes par un ensemble denzymes localisés dans les mitochondries et le REL. Les hormones diffusent ensuite librement à travers la membrane plasmique, du fait de leur caractère lipophile. 3) les neuromédiateurs : ils sagit principalement dacides aminés (glutamate par exemple), damines biogènes (c'est-à-dire des dacides aminés ayant subi une réaction enzymatique simple ; par exemple le tryptophane qui donne la sérotonine) ou de neuropeptides (endorphines par exemples). Les neuromédiateurs exercent principalement une fonction de neurotransmetteur et sont libérés par exocytose de vésicules synaptique. Il sagit donc dune action de type paracrine. B) Cellules épithéliales sécrétrices : Les cellules épithéliales sécrétrices peuvent se présenter sous 3 formes : i) il peut sagir de cellules isolées au sein dun épithélium de revêtement (ex cellules caliciformes = cellules à pôle muqueux ouvert), ii) il peut sagir de cellules formant un épithélium de revêtement (le seul exemple dans lorganisme est donné par lépithélium gastrique qui est un épithélium de revêtement sécrétoire) iii) il peut sagir dun assemblage de cellules réunies par du tissu conjonctif. On parle alors de glandes. Le produit de sécrétion des cellules épithéliales sécrétrices sexerce toujours localement dans une cavité ouverte de l'organisme ou à la surface de la peau. On parle de sécrétion exocrine (dans le milieu extérieur) par opposition aux sécrétions endocrines (dans le milieu intérieur). Le terme de glande correspond à toute structure anatomique regroupant des cellules sécrétrices, quelles soient épithéliales ou non. Cest pourquoi on parle de glandes endocrines bine quil ne sagisse pas dépithéliums glandulaire. Certaines glandes ont une fonction amphicrine cest-à-dire à la fois endocrine et exocrine. Le meilleur exemple est donné par le foie dont les cellules sécrétrices, le hépatocytes, synthétisent la bile (sécrétion exocrine déversée dans le tube digestif) et des hormones tel que l « Insulin Growth Factor » (IGF) qui sont déversées dans le sang. C) Epithéliums glandulaires : 1) définition : les épithéliums glandulaires au sens strict du terme répondent aux deux critères suivants : i) ces sont des glandes microscopiques ou des organes glandulaires formés de cellules épithéliales sécrétrices ; ii) leur développement requière l'invagination d'une structure épithéliale plane qui formera ultérieurement un épithélium de revêtement. Au cours du développement, on observe ainsi la séquence prolifération, bourgeonnement, puis invagination, sans rupture de continuité avec lépithélium dorigine. Ces deux critères sont remplis par l'ensemble des glandes exocrines. 2) classification a) critères morphologiques : les glandes exocrines possèdent une portion sécrétrice et dans la grande majorité des cas un canal excréteur formé d'un épithélium de revêtement qui exerce également des fonctions sécrétrices. Il faut toutefois savoir que la portion excrétrice est parfois absente. La classification des glandes exocrines est basée sur la morphologie du canal excréteur et de la portion sécrétrice des glandes exocrines. La portion sécrétrice peut adopter 4 formes : tubuleuse droite (en forme de tube droit), tubuleuse contourné (en forme de tube enroulé), acineuse (en forme de petite grappe de cellules constituant un acinus) ou alvéolaire (en forme de poire). b) critères fonctionnels : il existe deux critères de classification fonctionnelle qui sont : la nature du produit sécrété et le mode d'extrusion de ce produit. * en fonction de la nature du produit sécrété on distinguera les glandes séreuses et les glandes muqueuses. - les glandes séreuses ont une sécrétion de consistance aqueuse, exclusivement formée de protéines. Dans la majorité des cas ces protéines sont des enzymes actives ou des zymogènes (pro-enzymes). La grande fluidité du produit de sécrétion explique le faible diamètre du canal excréteur dont la lumière nest pas visible en microscopie optique (contrairement aux glandes muqueuses). Les cellules à sécrétion séreuse présentent une morphologie ultrastructurale spécifique caractérisée par : i) un pôle basal très riche en citernes de REG et en mitochondries ; ii) un noyau localisé au pôle basal de la cellule à distance du domaine basal de la membrane plasmique ; iii) un appareil de golgi très développé, toujours supranucléaire et dont la face de maturation est tournée vers le pôle apical ; iv) des grains de sécrétion, localisées au pôle apical de la cellule.
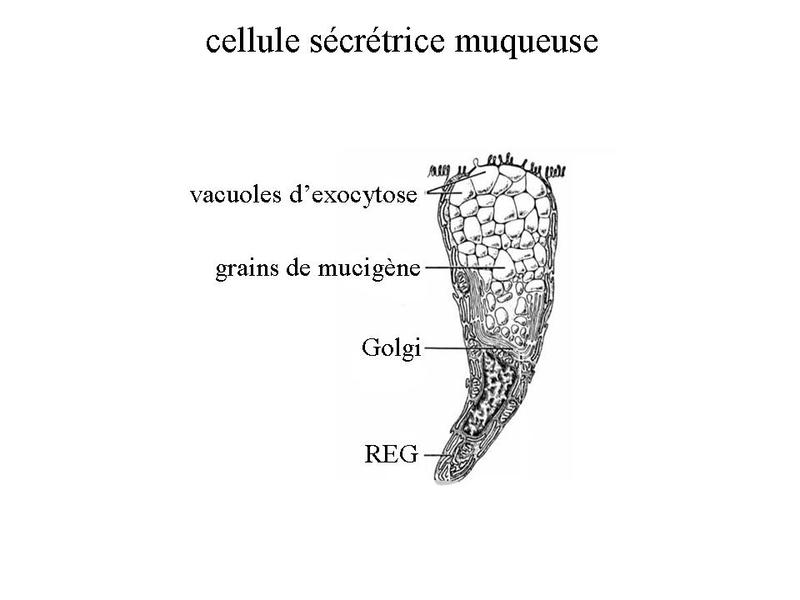
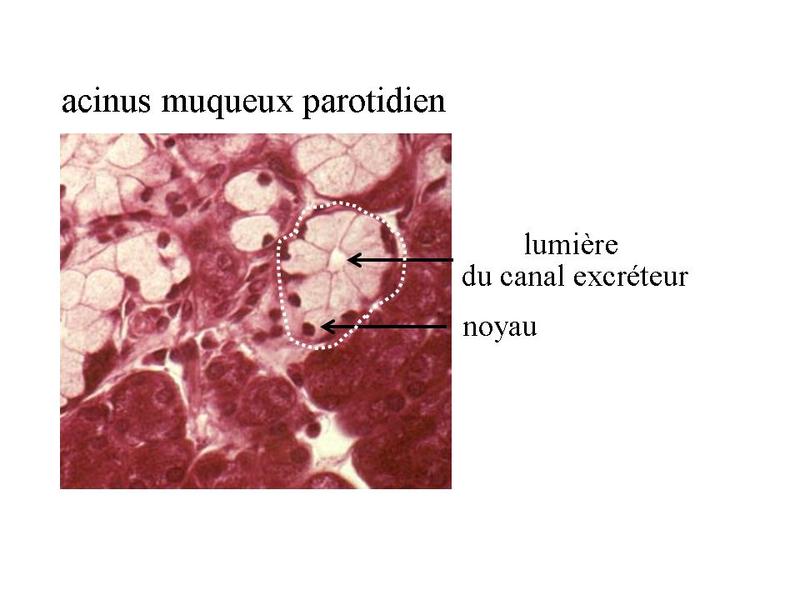
- les glandes muqueuses ont une sécrétion de consistance visqueuse nommée mucus et constituée de glucides et de glycoprotéines (dou la détection par la coloration PAS). La viscosité et la densité du produit de sécrétion expliquent labsence daffaissement du canal excréteur qui est visible en microscopie photonique. Les cellules à sécrétion muqueuse présentent certaines caractéristiques ultrastructurales : i) la présence dun nombre très important de grains de mucigènes et de vacuoles contenant du mucus mature, dans les deux tiers supérieurs du cytoplasme ; ii) le refoulement du noyau et des organites cellulaires à proximité du domaine basal de la membrane plasmique. Les glandes à sécrétion exclusivement muqueuse sont rares, ce type de sécrétion étant déjà largement produit par les cellules caliciformes disséminées dans les épithéliums de revêtement.
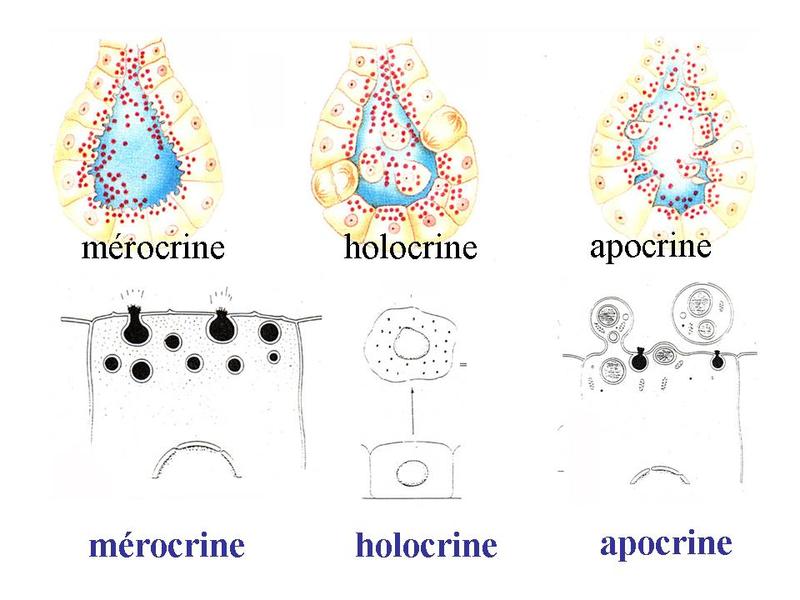
- les glandes séromuqueuses qui possèdent un contingent de cellules à sécrétion séreuse et un contingent de cellules à sécrétion muqueuse. Ce sont les plus fréquentes et le plus souvent elles réunissent des acinus purement muqueux, des acinus purement séreux et des acinus séro-muqueux (acinus réunissant des cellules à sécrétion muqueuse et des cellules à sécrétion séreuse). Exemple : les glandes sous-maxillaires qui participent à la formation de salive. Certains produits de sécrétion des glandes exocrines n'entrent dans aucune de ces catégories. C'est le cas du sébum sécrété par les glandes sébacées (contient essentiellement des lipides et des débris cellulaires) ou du lait sécrété par les glandes mammaires et qui contient des protéines et des lipides. * en fonction du mode d'extrusion du produit on distingue : - les glandes mérocrines ou l'extrusion se fait par exocytose (cas le plus fréquent, glandes sous-maxillaires) - les glandes holocrines ou l'ensemble de la cellule glandulaire est expulsée de la glande avec son produit de sécrétion (cas des glandes sébacées qui sécrètent le sébum) - les glandes apocrines ou le produit de sécrétion est sécrété avec une portion de la membrane plasmique apicale. La fraction lipidique du lait est excrétée de cette façon par la glande mammaire en lactation alors que la fraction protéique est libérée par mérocrinie.
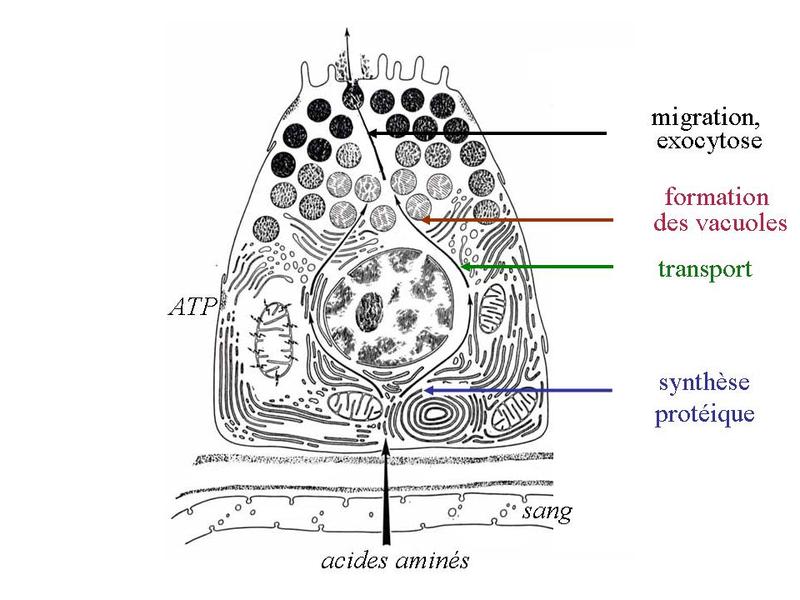
3) physiologie de la sécrétion épithéliale : La synthèse et lextrusion des produits de sécrétion suit plusieurs phases successives dont la nature, la durée et la régulation sont spécifiques du produit sécrété. Dans les glandes séreuses, ces phases sont les suivantes : i) captation et transport dacides aminés depuis la MEC à travers la membrane plasmique basale. Il sagit dun transport actif nécessitant la consommation de molécules dATP qui seront fournies par les mitochondries voisines ; ii) synthèse protéique au niveau du REG ; iii) transport des protéines vers lappareil de Golgi ; v) formation des vacuoles de sécrétions (grains de zymogènes) par bourgeonnement de la face trans de lappareil de Golgi ; vi) maturation, migration et libération des grains de zymogènes.
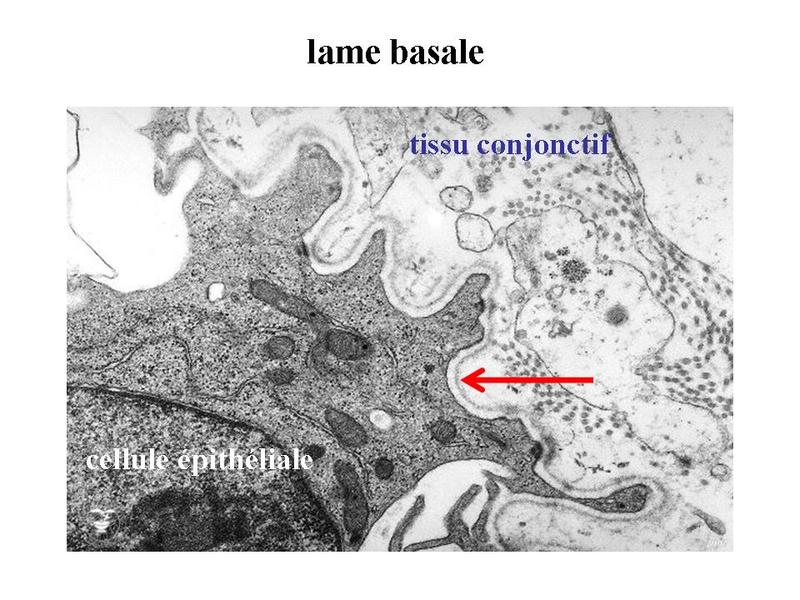
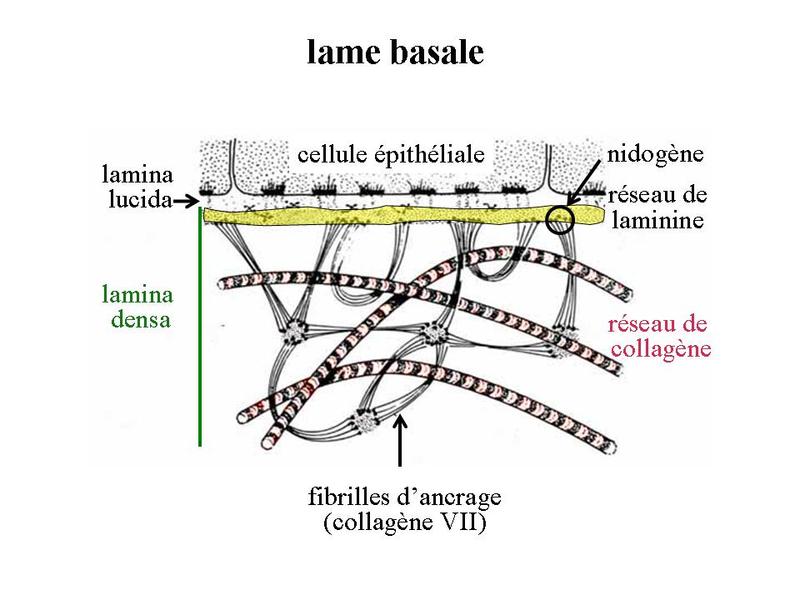
Dans les glandes muqueuses, ces phases sont les mêmes à la différence des phénomènes dajouts ou de modifications des résidus glycosylés au niveau du Golgi. Par ailleurs, à la différence des glandes muqueuses dont la sécrétion est le plus souvent continue et non régulée, les glandes séreuses sont soumises à une régulation fine de leur activité. Le contrôle de lactivité de synthèse et ou dexcrétion seffectue à 3 niveaux : i) niveau neuronal par lintermédiaire de synapses neuro-glandulaires ; ii) niveau hormonal via des récepteurs hormonaux spécifiques exprimés dans certaines glandes (vésicules séminales et prostate sous contrôle de la testostérone) ; iii) niveau musculaire par lintermédiaire de cellules à activité contractile, les cellules myoépithéliales qui bordent certains acinus séreux. V) LA LAME BASALE : 1) structure de la lame basale : la lame basale forme une couche complexe de molécules de la MEC établissant un contact étroit avec la membrane plasmique des cellules épithéliales. Une lame basale entoure également un certain nombre de cellules non épithéliales (cellules musculaires, cellules de Schwann, cellules endothéliales). La lame basale est visible en microscopie optique sous la forme d'un trait rouge après coloration au PAS. En microscopie électronique la lame basale prend la forme d'un fin feutrage de filaments irréguliers s'orientant dans les trois plans de l'espace. Au niveau des épithéliums, elle est constituée de deux couches superposées : la lamina rara (ou lamina lucida) qui est transparente aux électrons, la lamina densa qui contient des molécules de collagène IV.
La lame basale résulte ainsi de l'interaction de deux réseaux moléculaires : un réseau de collagène IV et un réseau de laminine.
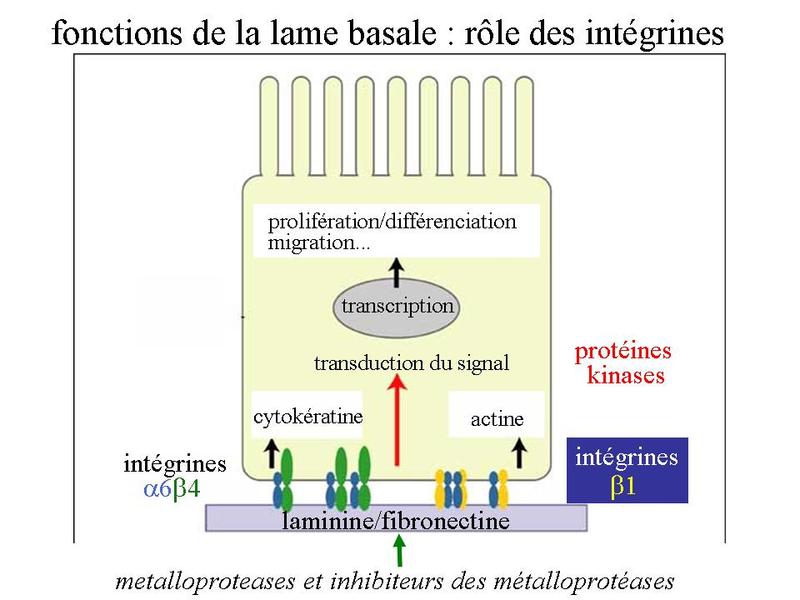
En regard de la lame basale, la surface cellulaire présente de nombreux récepteurs à des molécules de la MEC : les intégrines qui sont des récepteurs à la lamnine et à la fibronectine, la molécule CD44 qui est un récepteur à l'acide hyaluronique, des récepteurs à de nombreuses cytokines, facteurs de croissance et chimiokines dont la biodisponibilité est régulée par la lame basale. 2) fonctions de la lame basale : la lame basale joue premièrement un rôle d'ancrage des cellules épithéliales au tissu conjonctif sous-jacent. Elle exerce également des fonctions de barrière permettant le filtrage ou la rétention de molécules (cytokines, facteurs de croissance, chimiokines) dérivant du sang et de la MEC sous-jacente. La lame basale participe au maintien de la cytoarchitecture et en particulier de la polarité des épithéliums. Enfin, on considère également que la lame basale participe au contrôle de la différenciation, de la prolifération et de la migration des cellules souches épithéliales. L'essentiel des fonctions exercées par la lame basale est le fruit d'interactions entre molécules d'adhérence (fibronectine, laminine) et les intégrines. Les intégrines forment une famille d'hétérodimères constitués de sous-unités alpha et beta. En fonction des unités alpha et beta qui les constituent, les intégrines lient différentes molécules de la MEC et en particulier la fibronectine et la laminine. On considère que les intégrines forment la principale voie de communication entre cellule et matrice. Les intégrines sont toujours reliés aux microfilaments d'actine (pour mémoire : à l'exception de l'intégrine alpha 6-beta 4 qui entre dans la constitution des hémidesmosomes et est relié aux filaments intermédiaires). Les intégrines activées par leur ligand induisent une signalisation intracellulaire large via le cytosquelette et l'activation de protéines kinases. Cette signalisation permet de moduler l'expression d'un ensemble de gènes qui auront des effets sur de nombreuses fonctions cellulaires essentielles telles que la prolifération, la différenciation ou la migration. Enfin, la composition de la lame basale est contrôlée en permanence par un ensemble de protéases nommées métalloprotéases et leurs inhibiteurs qui au niveau des épithéliums sont essentiellement synthétisées par les principales cellules du tissu conjonctif sous-jacent : les fibroblastes.
|

Site créé avec ViaBloga
-- Modèle rdc par Jon Roobottom,
Paul Lloyd et
Stéphane Gigandet --
![]()