A la différence du tissu osseux mature, le tissu cartilagineux
mature présente un faible taux de renouvellement. Il ne
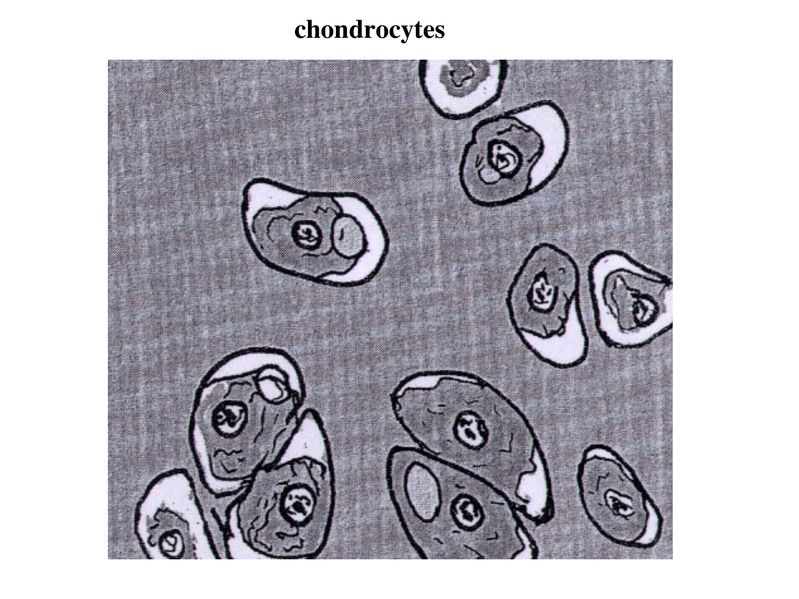
contient quun seul type cellulaire : les chondrocytes.
Les chondrocytes sont des cellules arrondies envoyant de
courts prolongements et qui sont entourées de matrice
cartilagineuse rigide mais non calcifiée. Cette matrice forme
des logettes anatomiques sans parois propres, les
chondroplastes.
Chaque chondroplaste contient un ou plusieurs chondrocytes. Les chondrocytes synthétisent la matrice cartilagineuse et interagissent avec elle via lexpression de récepteurs membranaires. Il sagit en particulier de la molécule CD44 dont le ligand est lacide hyaluronique et dIntégrines qui interagissent avec de nombreuses molécules matricielles. Les chondrocytes présentent les caractéristiques morphologiques de cellules à activité sécrétrice: le REG et lappareil de golgi sont développés. Cette activité de synthèse protéique correspond essentiellement à la sécrétion de lensemble des protéines de la matrice cartilagineuse. Toutefois, les chondrocytes synthétisent également à bas bruit des molécules induisant la résorption de la matrice cartilagineuse. Parmi ces molécules, citons les métalloprotéases ("matrix metalloprotéases : MMP"). Chez ladulte, cette activité de résorption est très faible en situation normale. Par ailleurs, les chondrocytes renferment des grains de glycogène et des vacuoles lipidiques dont les fonctions sont encore mal connues. Enfin, il faut noter que les chondrocytes du tissu cartilagineux mature sont considérés comme des cellules post-mitotiques.
2) la matrice : Chez l'adulte, on distingue 3 types de tissu
cartilagineux qui se distinguent essentiellement par
la composition de leur matrice.
Pour mémoire : Les chondrocytes sont des cellules post-mitotiques in vivo mais capables de proliférer in vitro après avoir été prélevées au niveau d'un cartilage mature. En conditions de culture standard, les chondrocytes adhèrent au plastique des boites de cultures et se dé-différencient rapidement en fibroblastes. Pour permettre leur prolifération et le maintien dun phénotype chondrocytaire, on doit cultiver les chondrocytes dans un gel contenant des facteurs de croissance et des molécules de la matrice cartilagineuse. Les chondrocytes ainsi générés peuvent être utilisés à des fins de recherche ou être réimplantés au sein de cartilages lésés. La greffe autologue de chondrocytes est actuellement utilisée pour le traitement de lésions cartilagineuse du sujet jeune. On a en effet observé que le potentiel chondrogènique des chondrocytes est étroitement dépendant de lage du donneur (plus faible à partir de 40 ans, très faible à partir de 70 ans). Par ailleurs, on considère que les chondrocytes forment une population hétérogène quant à leur capacité à proliférer in vitro. Au sein dun tissu cartilagineux articulaire, on observera ces différences fonctionnelles lorsquon compare chondrocytes superficiels et chondrocytes profonds.
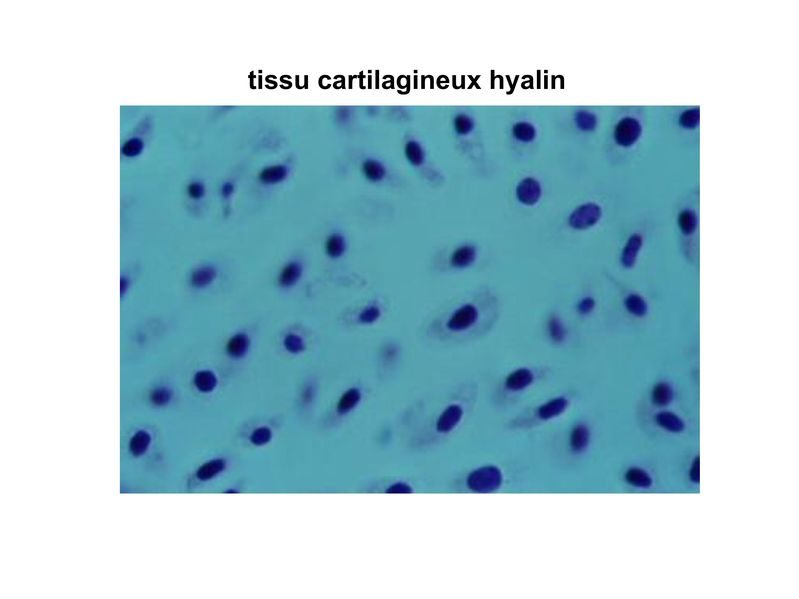
a) le tissu cartilagineux hyalin : c'est le plus répandu et il
est localisé au niveau des cartilages articulaires, des
cartilages trachéo-bronchiques ainsi qu'au niveau des
cartilages intercostaux. La matrice y contient différents
types de collagène mais majoritairement du collagène type
II. On considère que le collagène type II représente 80 à 90%
du contenu en collagène de la matrice du tissu cartilagineux
hyalin. Le collagène type II nest pas strictement spécifique des
tissu cartilagineux mais présent essentiellement dans ces
tissus.
Quelque soit sa localisation, le collagène type II s'organise en
microfibrilles et non pas en fibres et en faisceaux comme le collagène
I. De ce fait, il nest pas observable en microscopie optique à l'aide des
colorations standards permettant la visualisation des faisceaux de
collagène (trichrome de Masson, Jaune Safran).Cette caractéristique
associée à labsence de vaisseaux et de nerfs confère un
aspect amorphe à la matrice.
Par ailleurs, le turn-over de ce réseau de collagène est
quasiment nul dans le tissu cartilagineux hyalin mature (donc
faible capacité de réparation tissulaire). Comme l'ensemble des
cartilages, la matrice du tissu cartilagineux hyalin contient
essentiellement de l'eau (70 à 80% du poids hydraté). Cette
grande capacité de rétention de l'eau est liée à la présence
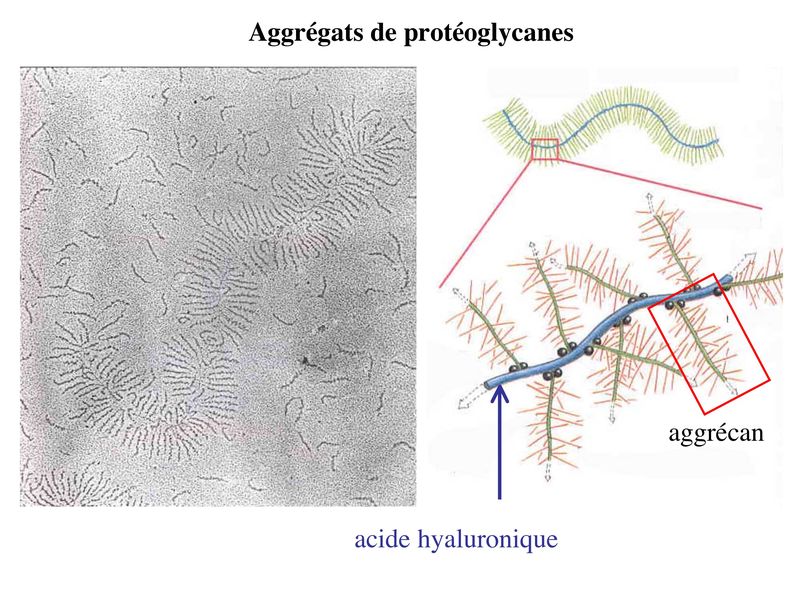
abondante d'aggrégats de protéoglycances contenant des
glycosaminoglycanes (GAG) sulfatés riches en radicaux
acides hydrophiles (pour mémoire : Ces GAG forment des
protéoglycanes sulfatés dont le principal est l'aggrécan).
(pour mémoire : Le métabolisme des protéoglycanes au sein du cartilage articulaire dépend des facteurs biomécaniques auxquels est soumise l'articulation. Lors d'une immobilisation complète, on observera une diminution de leur synthèse avec un amincissement cartilagineux associé à une fuite d'eau tissulaire. Par ailleurs, la fonction de rétention deau exercée par les protéoglycanes est responsable la déformabilité et de la résistance du tissu cartilagineux hyalin. Ainsi, lorsqu'une forte pression est exercée sur un cartilage articulaire par exemple, une partie de l'eau est expulsée mais l'augmentation de la pression osmotique induit un afflux d'eau qui rétablit la concentration en eau et une forte pression hydrostatique.
Outre la rétention de l'eau, le rôle des protéoglycanes de la
matrice cartilagineuse est de permettre la diffusion et/ou la
fixation de nombreuses molécules (facteurs de croissance,
cytokines, métabolites, nutriments, hormones...) nécessaires à
la fonction des chondrocytes. Dans le cas des cartilages
articulaires, le tissu cartilagineux est nourri par des molécules
diffusant à partir du liquide synovial et du tissu osseux
sous-jacent (os sous-chonral). Dans les autres types de
cartilages hyalin, comme par exemple le carttilage trachéo-
bronchique) ces molécules dérivent dun tissu conjonctif
vascularisé entourant le tissu cartilagineux : le périchondre.
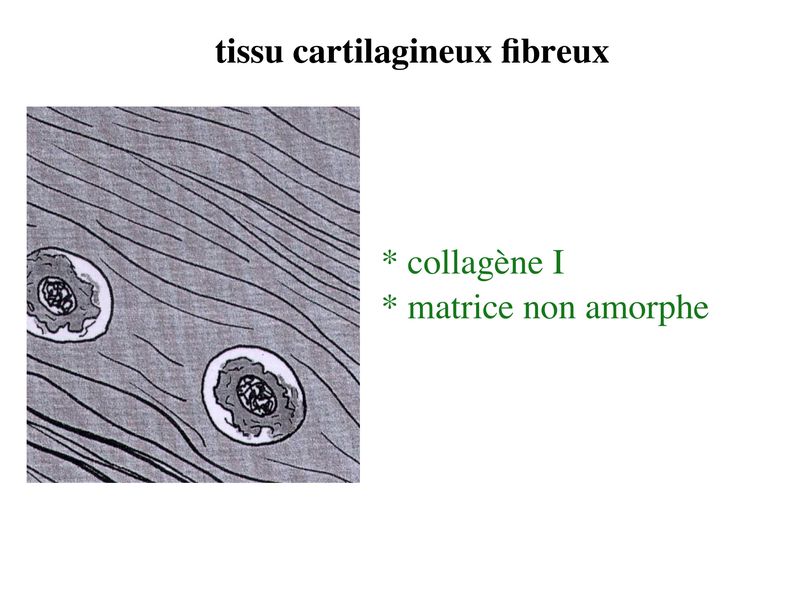
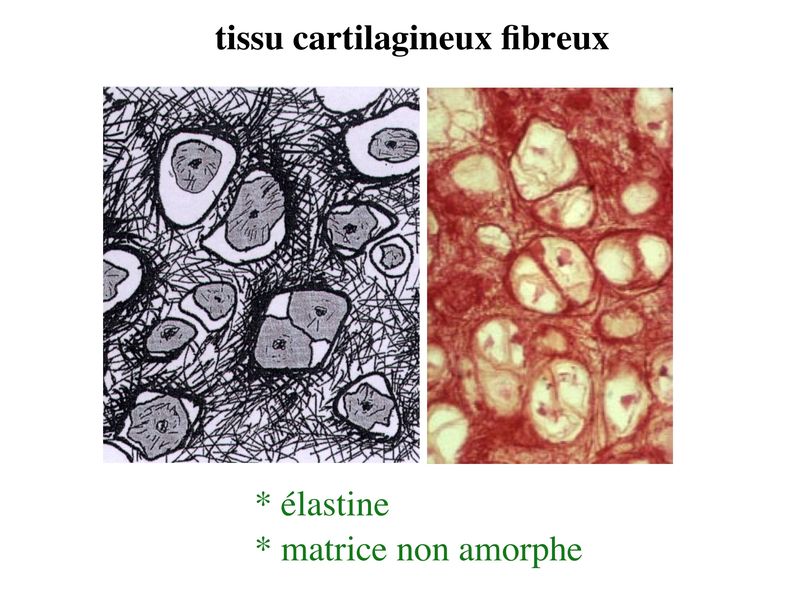
b) le tissu cartilagineux fibreux : encore nommé
fibrocartilage, il diffère du cartilage hyalin par la présence
abondante de fibres de collagène type I. Ces fibres forment
des faisceaux orientés détectables en microscopie optique
après coloration au trichrome de Masson (ou au Jaune
Safran).
Par conséquent la matrice de ce tissu cartilagineux ne peut être
considérée comme amorphe. Le fibrocartilage est présent au
niveau du tendon d'Achille (plus exactement au niveau du
point d'insertion du tendon d'Achille), des ménisques et des
disques intervertébraux.
c) le cartilage élastique : le cartilage élastique est
caractérisé par l'abondance des chondrocytes et par la
présence d'un réseau tridimensionnel de fibres élastiques
(fibres que l'on peut détecter par une coloration à l'orcéine).
Il est localisé essentiellement au niveau des ailes du nez, des
pavillons de l'oreille et de l'épiglotte.
II) le tissu cartilagineux immature : :il est essentiellement
observé chez le foetus mais également chez lenfant et
ladolescent ou il participe à la croissance osseuse
post-natale.
1) les cellules : contrairement au tissu cartilagineux mature,
le tissu cartilagineux immature contient des cellules à riche
activité mitotique. Selon le mode de croissance cartilagineuse, il
peut sagir de chondroblastes (précurseurs des chondrocytes)
ou de chondrocytes eux-mêmes.
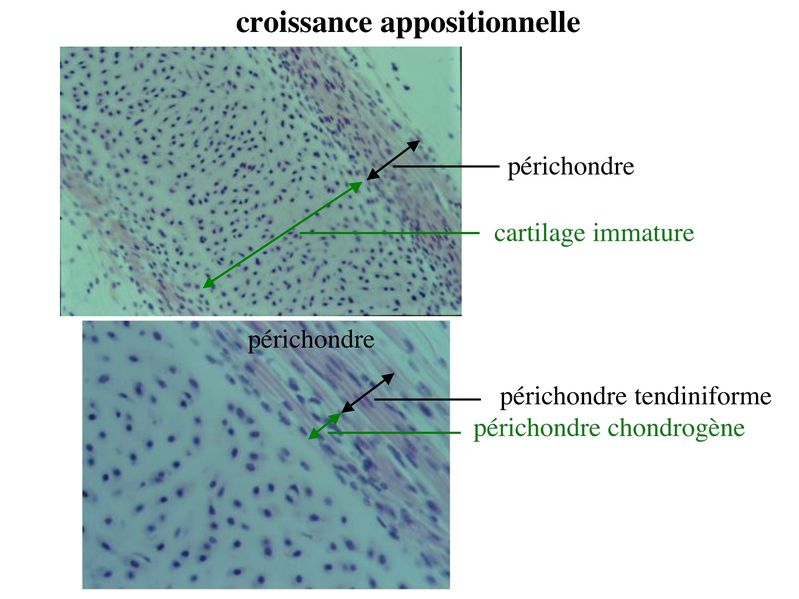
a) la croissance dite appositionnelle(ou périchondrale) :
sopère par prolifération puis différenciation de chondroblastes
en chondrocytes. Dans ce cas, les chondroblastes dérivent de
cellules souches mésenchymateuses localisées dans la couche
la plus interne du périchondre, le périchondre chondrogène.
Celui-ci forme un tissu conjonctif lâche (pauvre en fibres et
richement vascularisé) qui se distingue de la couche externe du
périchondre nommée couche tendiniforme qui est formée de
tissu conjonctif dense. La croissance appositionnelle est
essentiellement observée au cours du développement ftal.
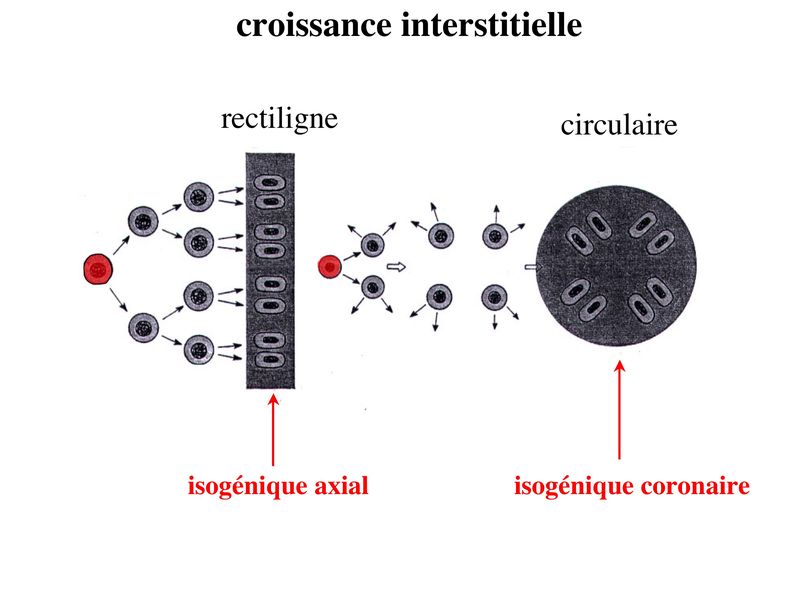
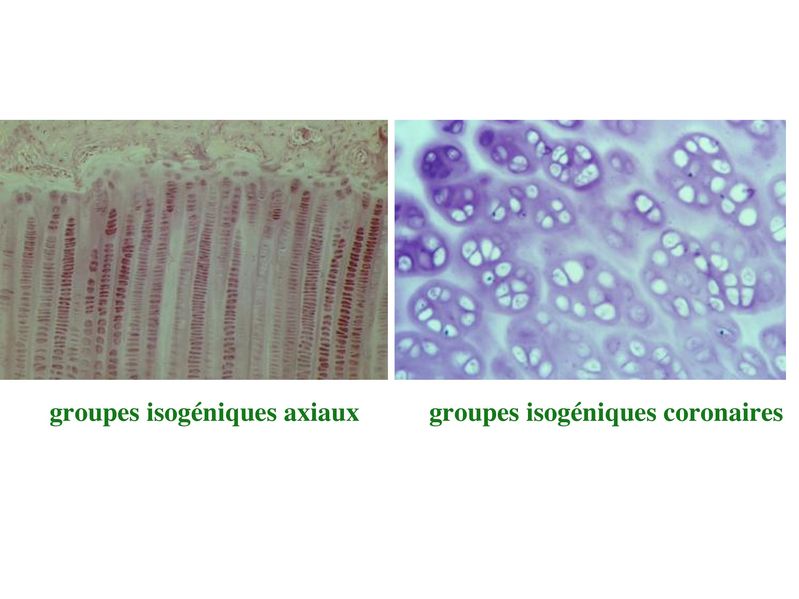
b) la croissance interstitielle : elle seffectue par mitoses
successives des chondrocytes eux-mêmes. Dans ce cas, les
cellules filles dun même clône cellulaire séloignent de la cellule
mère en se disposant soit de manière rectiligne soit de manière
circulaire. Elles forment ce quon appelle des groupes
isogéniques axiaux ou coronaires. Ces différentes
dispositions permettent la croissance axiale ou
circonférentielle du cartilage.
Durant ce processus on observe souvent a présence de
plusieurs chondrocytes au sein du même chondroplaste.
La croissance interstitielle est observée chez le foetus mais
également au cours de la croissance osseuse post-natale
des os long. En effet jusqu'à la puberté, la métaphyse des os
longs renferme un cartilage de croissance nommé cartilage
de conjugaison qui participe à la croissance osseuse.
2) la matrice : La matrice du tissu cartilagineux immature est
proche de la matrice du cartilage mature hyalin mais présente
un certain nombre de spécificités. On retient en particulier la
présence non négligeable de collagène IX.
III) Tissu cartilagineux et développement osseux le
développement du tissu osseux s'effectue schématiquement en
2 phases qui sont :
i) l'ossification primaire c'est-à-dire la formation initiale du
tissu osseux chez le foetus,
ii) lossification secondaire cest-à-dire la croissance et la
maturation osseuse qui s'achèvent à la fin de la puberté.
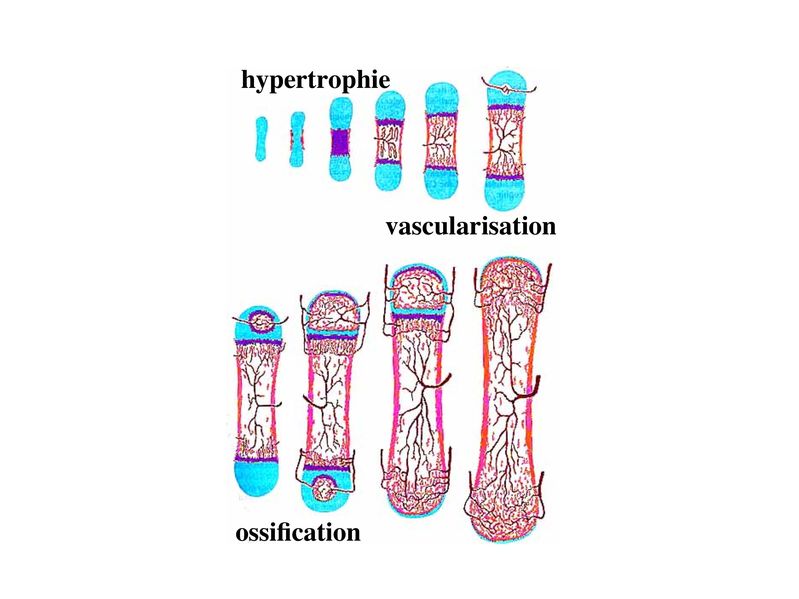
1) tissu cartilagineux et ossification primaire : on
désigne sous le terme d'ossification endochondrale la
transformation d'un tissu cartilagineux hyalin en tissu osseux.
Ce type dossification est à distinguer de lossification dite
endoconjonctive qui consiste en la transformation osseuse dun
tissu conjonctif non spécialisé (Pour mémoire : cette
ossification endoconjonctive est observée au niveau du périoste
qui entoure la diaphyse des os long ; on parle dossification
périostique). Dans les os longs, l'ossification primaire
endochondrale débute au centre de la diaphyse par un
processus d'hypertrophie des chondrocytes. Celui-ci
s'accompagne de 3 évènements :
i) l'élargissement des chondroplastes au dépends de la
MEC
ii) la synthèse de facteurs angiogéniques dont le VEGF
("vascular endothelial growth factor") induisant la
vascularisation locale du tissu cartilagineux
iii) la calcification du tissu cartilagineux (calcification
n'équivaut pas à ossification).
Puis, la phase initiale dhypertrophie des chondrocytes sachève
par leur mort cellulaire programmée (apoptose). Les
chondroplastes élargis sont donc vidés de leur contenu et vont
devenir confluents. Ils sont alors séparés par des travées de
tissu cartilagineux calcifié et vascularisé. Les vaisseaux
sanguins pénètrent alors dans les chondroplastes confluents et
permettent l'arrivée de cellules souches
mésenchymateuses circulantes qui vont générer des
ostéoblastes. Les ostéoblastes permettent l'ossification des
travées de tissu cartilagineux et conduisent à la formation d'un
point d'ossification primaire au centre de la diaphyse. Par la
suite, on observera de proche en proche le recrutement et la
prolifération de nouveaux chondrocytes entourant le front
dossification.
Dans les os longs, lorsque l'ossification primaire de la diaphyse
est suffisamment avancée, débute l'ossification primaire des
épiphyses. Il s'agit également d'une ossification
endochondrale évoluant de manière centrifuge à partir d'un
point d'ossification centro-épiphysaire. Cette ossification
préserve une zone de cartilage articulaire en regard de la
cavité synoviale et une zone de cartilage immature (le
cartilage de conjugaison) au niveau de la métaphyse.
L'ensemble du processus d'ossification primaire ne conduit en
fait qu'à lossification de travées de tissu cartilagineux : on
parle d'os primaire endochondral (os immature, non lamellaire,
Cf. cours tissu osseux). Lors d'une deuxième phase qui s'étend
jusqu'à la fin de la puberté se déroule l'ossification secondaire
c'est-à-dire d'une part le remplacement complet de cet os
immature par un os mature dit lamellaire, et d'autre part la
croissance en longueur et en épaisseur des os.
(pour mémoire : tout phénomène de réparation osseuse, après fracture par exemple, saccompagne dune récapitulation des phénomènes caractérisant lossification endochondrale. On observe ainsi la formation initiale dun modèle cartilagineux au sein duquel les chondrocytes dérivent de cellules souches mésenchymateuses. Ce modèle cartilagineux se calcifie et est progressivement remplacé par du tissu osseux).
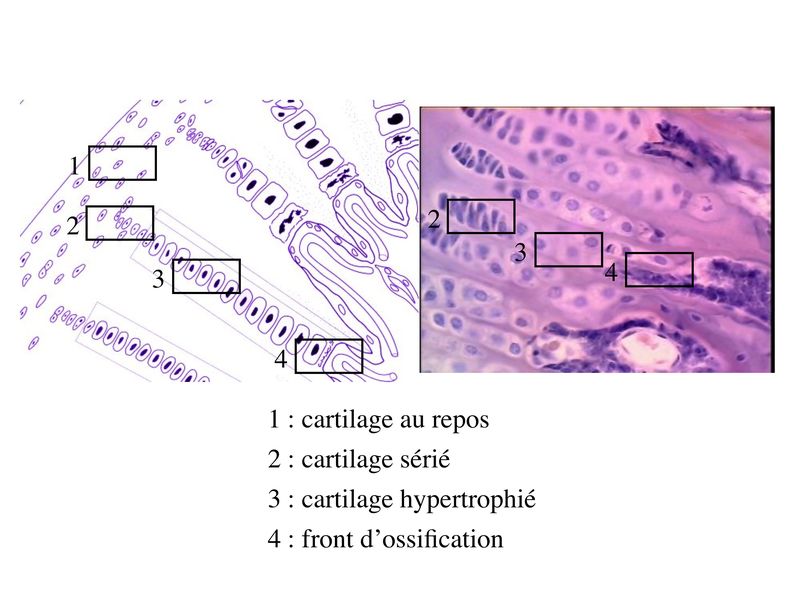
2) tissu cartilagineux et ossification secondaire : la
croissance en longueur des os est de type endochondral et fait
intervenir les cartilages de conjugaison. Le cartilage de
conjugaison est organisé en colonnes et en couches
successives qui sont individualisables en microscopie optique.
La zone du cartilage hyalin la plus éloignée du front
dossification constitue une réserve de chondrocytes au
repos. Puis on trouve une zone de cartilage sérié ou les
chondrocytes prolifèrent et saplatissent.
Enfin, à proximité du front dossification se trouve une zone
dite de cartilage hypertrophié ou les chondrocytes
augmentent leur volume par un facteur de 5 à 10. Ces
chondrocytes hypertrophiés vont exercer les fonctions
suivantes :
i) calcification de la matrice cartilagineuse,
ii) vascularisation du tissu cartilagineux,
iii) recrutement puis différenciation ostéoblastique de
cellules souches mésenchymateuses. On considère que ces
cellules souches proviennent du tissu conjonctif non spécialisé
associé au tissu osseux (endoste, périoste, tissu conjonctif
associé à la moelle osseuse rouge).
Pour méméoire : La croissance en épaisseur des os longs nest pas de type endochondral et se fait par apposition de couches successives de lamelles osseuses à partir du périoste. Il s'agit d'une croissance endoconjonctive au cours de laquelle on observe la différenciation de cellules souches mésenchymateuses en ostéoblastes, sans intervention dun autre type cellulaire et sans migration par voie sanguine des cellules souches mésenchymateuses.
3) contrôle moléculaire de l'ossification endochondrale :
a) facteurs systémiques : pendant toute la période de
croissance post-natale, la croissance en longueur des os longs
est sous la dépendance de facteurs hormonaux agissant
essentiellement sur les cartilages de conjugaison. Il sagit
principalement de lIGF1 (dont lhormone de croissance GH
stimule la production par le foie) et des hormones sexuelles
i.e. les oestrogènes chez la femme et les androgènes chez
lhomme. Ce contrôle hormonal explique la poussée de
croissance osseuse au moment de la puberté. Il faut noter que
les chondrocytes nexpriment que des récepteurs aux
oestrogènes et non aux androgènes. Leffet des androgènes
chez le garçon nécessite laction dune enzyme intracellulaire
qui métabolise les androgènes en oestrogènes. À lissue de
cette période, le stock de chondrocytes contenus dans le
cartilage de conjugaison est épuisé, le cartilage de conjugaison
est totalement remplacé par du tissu osseux et la taille
définitive de lindividu est atteinte.
b) facteurs locaux : les chondrocytes hypertrophiés
synthétisent un ensemble de molécules qui permettent : i)
soitlossification du cartilagineux immature au cours de
lossification primaire, ii) soit lossification des cartilages de
croissance au cours de lossification secondaire.
i) le VEGF : le « vascular endothelial growth factor » est
un puissant agent angiogénique permettant la formation de
vaisseaux (on parle de néovascularisation) dans la zone des
chondrocytes hypertrophiés.
Cette vascularisation permet i) un soutien
nutritif et métabolique ii) le contrôle moléculire sytémique par les facteurs
solubles sanguins décrits plus haut, iii) la lapport de cellules souches
mésenchymateuses sanguines (uniquement chez le foetus). Par ailleurs, le
VEGF exerce une activité chimiotactique (cest-à-dire une
activité dinduction de la migration) sur les ostéoblastes,
permettant ainsi de diriger leur migration vers la zone
d'ossification.
ii) la molécule Indian Hedgehog (IHH) : il sagit dun
polypeptide synthétisé par les chondrocytes hypertrophique ou
pré-hypertrophiques. Indian Hedgehog agit sur 3 phases
essentielles de lossification :
i) la prolifération des chondrocytes,
ii) leur différenciation en chondrocytes hypertrophiés,
iii) la maturation des ostéoblastes.
(pour mémoire : Il existe une pathologie humaine, nommée brachydactylie, qui est liée à une mutation de lIHH. Cette pathologie est caractérisée par la longueur anormalement faible des phalanges et des métacarpes).
IV) pathologies du tissu cartilagineux :
1) pathologies mécaniques et dégénératives : elles
concernent essentiellement les cartilages articulaires et les
fibrocartilages.
a) larthrose Elle peut toucher de nombreuses articulations mais en particulier la hanche (coxarthroes), le genou (gonarthrose) et la colonne vertébrale. Elle se traduit par : 1) lamincissement puis la disprition du cartilage (la disparition du cartilage saccompagne au plan radiologique par la disparition de linterligne articulaire); 2) une densification réactionnelle et des micro-fractures de l'os sous-chondral 3) la formation dexcroissances ostéocartilagineuses nommées ostéophytes . Puis on observe la disparition complète du cartilage et la survenue de lésions osseuses par frottement. La coxarthrose primitive survient le plus souvent entre 50 et 70 ans et est considérée comme dorigine multifactorielle (facteur héréditaires + facteurs aggravants tel que lobésité et les microtraumatismes). L'arthrose secondaire survient plus précocement chez des sujets aux antécédents de traumatisme, danomalies de croissance ou d'anomalies de développement tel que la luxation bilatérale de hanche (pathologie actuellement détectable et traitable à la naissance).
b) lésions des fibrocartilages : latteinte dégénérative des
fibrocartilages est fréquente au-delà de 50 ans. On peut
observer en particulier une dégénérescence des disques